Results
Response at the fundamental frequency included all the responses
measured at the first harmonic, but the converse was not always true. When
multiplication by the cosine-bell window was included as a step in the
processing, harmonic responses mimicked responses at the fundamental frequency.
When the window was omitted, however, harmonic responses were less robust, and
comparisons did not always attain significance in intervals corresponding to
periods of significance at the fundamental frequency. Because the harmonic
data added nothing to the data obtained at the fundamental frequency, only the
latter are reported here.
The most consistent electrophysiological responses occurred at the two lateral
derivations (figure 2). Note the difference between these data and
conventional ERP recordings. The graphs are not simply the averaged signal
versus time. Rather, the vertical deflection of each trace measures the amount
of EEG power at the 8.9Hz frequency of interest. Each graph contains two
traces, one (solid) representing the response to left-field targets, and the
other (dotted) representing the response to right-field targets. The
statistical test was applied not to either of these traces by itself but rather
to the difference between the two traces.
During the first 300ms following the stimulus, the right hemisphere responded
with an increase in phase-locked amplitude both to left-field targets and to
right-field targets, whereas the left hemisphere responded significantly more
to right-field targets than to left-field targets (sign test, p<0.04
in the latency ranges 161ms-283ms poststimulus in bin 0, 98ms-203ms
poststimulus in bin 1, 66ms-87ms poststimulus in bin 2, and 45ms-87ms
poststimulus in bin 3). The latency of this differential response in the left
hemisphere decreased with increasing intertarget intervals. (At 308ms-318ms
poststimulus, on the falling edge of this response, Friedman's test
 23=0.3, p~0.04; post hoc
21=0, p~0 between bins 0 and 1. At 178ms
poststimulus, on the rising edge, Friedman's test 23=0.3,
p~0.04; post hoc 21=0, p~0 between bins
0 and 2 and between bins 0 and 3.) It is important to distinguish between
response latencies and intertarget intervals. In this discussion, the term
`latency' refers to the interval of time that separates a target and an
electrophysiological response to that target, whereas the term `intertarget
interval' refers to the interval of time that separates that same target from
the previous target.
23=0.3, p~0.04; post hoc
21=0, p~0 between bins 0 and 1. At 178ms
poststimulus, on the rising edge, Friedman's test 23=0.3,
p~0.04; post hoc 21=0, p~0 between bins
0 and 2 and between bins 0 and 3.) It is important to distinguish between
response latencies and intertarget intervals. In this discussion, the term
`latency' refers to the interval of time that separates a target and an
electrophysiological response to that target, whereas the term `intertarget
interval' refers to the interval of time that separates that same target from
the previous target.
At longer latencies, amplitudes in each hemisphere tended to be greater in
response to ipsilateral targets (that is, cues to shift attention to the
contralateral visual hemifield) than to contralateral targets (that is, cues to
shift attention to the ipsilateral visual hemifield), but there was an
important exception to this generalisation in the right hemisphere at long
intertarget intervals. In bin 0, phase-locked amplitude in the right
hemisphere was greater in response to right targets than to left targets in the
latency ranges 728ms-784ms, 920ms-931ms, and 1340ms-1368ms (sign test,
p<0.04). In bin 1, however, this right-hemisphere effect inverted,
so that phase-locked response was greater to left targets than to right targets
in the latency range 805ms-836ms (sign test, p<0.04). In bins 2 and
3, the effect did not attain significance in either direction.
A similar effect manifested in the left hemisphere, but without the inversion
in bin 1 and disappearance in bin 2. In bin 0, left-hemisphere phase-locked
amplitude in response to left-field targets exceeded response to right-field
targets in the latency range 1239ms-1442ms (sign test, p<0.04). In
bin 1, the same effect occurred, but in the lesser latency range 889ms-903ms
(sign test, p<0.04). This progression to earlier latencies continued
in bin 2, at 623ms-672ms (sign test, p<0.04), and the effect
disappeared in bin 3. This progression must be viewed merely as a trend,
though, since Friedman tests in these intervals did not reach significance.
Tests of non-phase-locked amplitudes revealed effects complementary to those
involving phase-locked amplitudes (figure 3). Since brain oscillations whose
phases do not have a constant relationship to the time of stimulus presentation
tend to average to zero (assuming a uniform distribution of phases), switching
the order of the amplitude computation and the averaging operation reveals
information about background activity. Whereas the phase-locked amplitudes
tended to become greater ipsilateral to the target (that is, contralateral to
the cued hemifield), non-phase-locked amplitudes tended to diminish
ipsilaterally, following a brief contralateral decrease.
At the lateral derivation in the right hemisphere, non-phase-locked activity
initially decreased more in response to left-field targets than in response to
right-field targets (sign test, p<0.04 at 150ms-262ms poststimulus in
bin 0, 112ms-182ms in bin 1, 126ms-248ms and 399-469ms in bin 2, and
164ms-360ms in bin 3). The latency of this response reached a minimum in bin 1
and became maximal in bin 3 (Friedman's test, 23=0.3,
p=0.04). Post hoc comparisons indicated significant differences
on the rising edge of this response (bins (0,2) at 136ms-147ms), and on the
falling edge early (bins (0, 2), (0, 3), (1, 3) at 262ms-287ms) and late (bins
(0, 3), (1, 3) at 325ms-339ms). At longer latencies, right-hemisphere
non-phase-locked amplitude was more suppressed in response to right-field
targets than to left-field targets (sign test, p<0.04 in the latency
ranges 476ms-500ms and 1004ms-1123ms in bin 0, 1078ms-1169ms in bin 1,
913ms-973ms and 1641ms-1725ms in bin 2, and 644ms-679ms in bin 3).
At the medial derivation in the right hemisphere, this pattern was inverted,
and somewhat less robust. In bin 0, no significant differences between
amplitudes in response to left targets and amplitudes in response to right
targets occurred until late in the epoch (sign test, p<0.04 from
1788ms to 1928ms). In bin 3, amplitude in response to right targets was less
than amplitude in response to left targets both initially and later in the
epoch (sign test, p<0.04 at 420ms-444ms, 581ms-647ms, 721ms-738ms,
and 1652ms-1659ms). In other bins, amplitude initially was more depressed in
response to right targets than in response to left targets (sign test,
p<0.04 in latency ranges 87ms-108ms and 283ms-315ms in bin 1, and
119ms-157ms in bin 2). At longer latencies, amplitude was more depressed in
response to left targets than in response to right targets (sign test,
p<0.04 at 1788ms-1928ms in bin 0 and at 980ms-1186ms in bin 2).
At the lateral derivation in the left hemisphere, the pattern appeared
complementary to that at the right-hemisphere lateral derivation.
Non-phase-locked activity initially decreased more in response to right-field
targets than to left-field targets (sign test, p<0.04 at 563ms-577ms
in bin 0 and 493ms-546ms in bin 3, with a trend at p=0.15 at 329ms-364ms
in bin 1). At longer latencies, non-phase-locked amplitude decreased more in
response to left-field targets than to right-field targets (sign test,
p<0.04 at 1883ms-1935ms in bin 0, 1379ms-1498ms in bin 2, and
861ms-871ms and 1179ms-1232ms in bin 3).
At the medial derivation, the pattern was again inverted. Amplitude initially
was more depressed in response to left targets than in response to right
targets (sign test, p<0.04 at 623ms-626ms in bin 0, 164ms-241ms in
bin 2, and 164ms-192ms in bin 3). At long latencies amplitude was more
depressed in response to right targets than in response to left targets (sign
test, p<0.04 at 752ms-787ms in bin 0, 654ms-731ms in bin 2, and
1746ms-1778ms in bin 3).
These results are complicated by the occurrence of small but systematic
deviations in eye position toward the attended locations (figure 4). Amplitude
changes in the horizontal electrooculogram indicate that eye movements were on
the order of one degree, nowhere near the distance between the targets, but
perhaps enough to influence the SSVEP. In a task that requires rapid,
consecutive shifts of attention, it seems, it is impossible to inhibit saccades
completely.
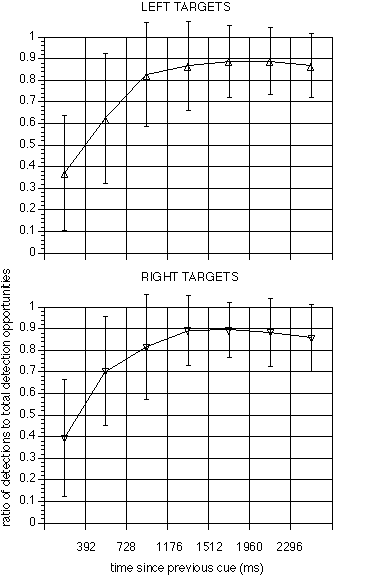
In the behavioural data, there was a highly significant effect of inter-target
interval on the ratio of correct target detections to total number of detection
opportunities (F(6, 66) = 43.27, p < 0.0001, figure 6).
Detection ratios increased rapidly through the lowest inter-target intervals,
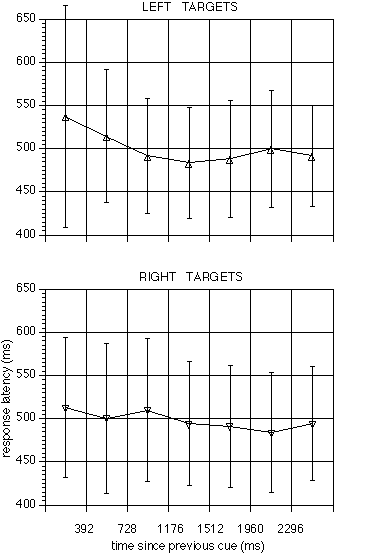
and peaked in the neighbourhood of 1500ms. Response latencies decreased
correspondingly (F(6, 66) = 3.49, p = 0.0047, figure 7).
Discussion
{kind=link}
{kind=link}
{kind=link}